摘要:功能性状和系统发育多样性都会影响生态系统功能,但在自然系统中,这些因素中哪一个最重要仍然知之甚少。利用西非森林和热带稀树草原的数据分析了(i)系统发育多样性如何补充解释地上碳(AGC)的功能性状属性;(ii)系统发育多样性与系统发育信号梯度上的功能性状属性相关;(iii)系统发育多样性和功能性状属性之间的途径将AGC与土壤和气候联系起来。系统发育多样性以平均成对距离(sesMPD)和平均最近分类单元距离(sesMNTD)的标准化效应大小来衡量。计算了与叶经济谱和植物生活史相关的四个性状的功能分散度(FDiss)和群落加权平均值(CWM)。
基于功能特征的模型解释了11%的AGC变异性。由于4个性状中有2个在系统发育上是保守的,在模型中加入系统发育多样性使AGC的解释方差增加了15%。与CWM相比,FDiss的系统发育多样性与性状关系的斜率对性状保守性的反应更敏感。植株最大高度的sesMPD和CWM对AGC有正向影响。反过来,植物最大高度的CWM随着土壤氮和气候湿度的升高而增加,而sesMPD与气候湿度负相关。虽然FDiss与sesMPD呈正相关,但在影响AGC与土壤氮和气候湿度并在其关联方面,其重要性不如sesMPD和植物最大高度的CWM。
结果表明,系统发育多样性对于AGC很重要,但并不能完全反映与群落广泛性状均值相关的功能机制。该研究还证明了环境在调节AGC中的作用,这种调控通过高植物高度驱动的群落适应度差异以及在干旱较少的环境中维持密切相关物种的进化过程来实现。
关键词:功能分散、最大株高、平均成对距离、系统发育信号、乔木和灌木稀树草原、西非
1.引言
为了解决这些问题,我们重点研究了西非的植被,众所周知,西非的植被在其地貌方面极为多样化,其中包括几内亚沿海半常绿植被、干旱森林、林地、乔灌木草原和草本草原。我们使用了在半干旱过渡区的森林--稀树草原马赛克中建立的246个森林样地收集的数据,这些森林样地横跨四个西非国家。森林--稀树草原是非洲特别重要的生物群落,是多种植物物种的宿主,提供重要的生态系统服务并在碳收支中发挥重要作用(Lykke等,2009)。然而,在生物多样性对EF影响的背景下,它们的研究在很大程度上还不够充分;并添加到我们在多个国家的可靠地块数据中,它们为这项研究提供了合适的背景,为我们对树木群落的理解做出了独特的贡献。基于与叶片、种子和木材特性相关的功能性状,我们计算了群落加权平均值(以下简称CWM)和作为功能多样性(以下简称FD)属性的功能分散度(FDiss)。我们利用了更新后的GBOTB的包含大于70,000个物种的宏系统发育树。扩展的巨型树(Smith和Brown,2018;Zanneetal,2014)构建了所有物种的扎根和时间校准的系统发育(图A1),在此基础上我们计算了MPD(sesMPD)和平均最近分类距离(sesMNTD)作为PD属性的标准化效应大小(图A2)。
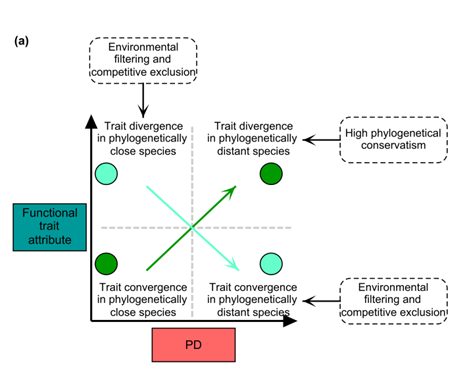
图1(a)
系统发育多样性(PD)与功能性状属性(FD和CWM)之间关系的概念框架
在(a)中,我们预计,在性状保守性较高的假设下,系统发育距离较远的物种群落表现出更大的功能分化,这将允许物种共存并在资源方面占据不同的生态位;然而,在强大的环境过滤下,共享相同非生物环境的共生物种预计会比预期更加相似(性状聚类;Caver-Bares等人,2004年)。
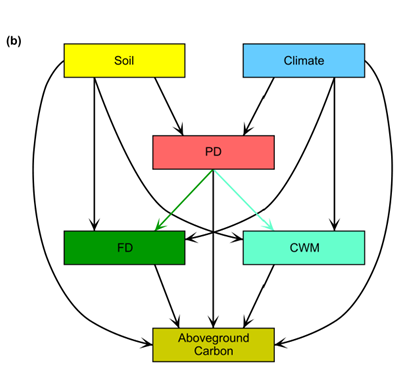
图1(b)
结构方程模型,测试PD、FD和CWM之间的相互关联途径如何将环境变量(土壤和气候)与地上碳(AGC)联系起来。
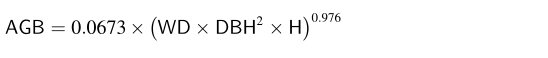
在(b)中,我们预计PD的增加通常与FD的增加有关,这反过来又会促进AGC,假设不同物种通常具有不同的资源需求和使用策略,从而使它们能够更好地分配资源并减少竞争。我们还预计环境条件会通过环境过滤和选择影响PD和功能性状属性(FD和CWM),而较好的环境条件(如高水分和养分有效性)会通过PD或功能性状属性(FD和CWM)促进AGC。
2.材料与方法
2.1.研究区域
研究区域位于西非半干旱的稀树草原和森林中,并覆盖了布基纳法索南部、加纳北部、多哥北部和贝宁北部的部分地区(图2)。
图2
地图显示了西非研究区的位置和样地分布

2.2.库存图及数据
我们使用了一个包括246个地块的广泛数据集,平均地块面积约为0.12ha。这些地块在地理上分布在贝宁北部(84个地块)、多哥北部(109个地块)、加纳北部(27个地块)和布基纳法索南部(26个地块),从而涵盖了广泛的环境变化。
在246个地块中,74个地块有火灾历史,并从数据集中删除。结果表明,共有172个样地和124个物种可供进一步分析。
2.3.特征数据
我们选择的功能性状涵盖了叶片和木材经济性状谱的相当一部分,同时也考虑了种子和树的大小:植株最大高度(Hm;m),木材密度(WD;g/cm3),比叶面积(SLA;mm2/mg),叶片干物质含量(LDMC;mg/g)和种子干质量(SDM;g)。
2.4.环境数据
本研究使用的环境数据包括3个气候变量和5个土壤变量。作为气候变量,我们使用了气温季节性、降水季节性和气候湿度指数。我们从CHELSA(Climatol--ogies在高分辨率的地球陆地表面地区;https://)数据库(Karger等,2017)。我们从非洲土壤剖面数据库(https://)检索了250m空间分辨率的土壤物理和化学性质信息。这些指标包括全氮(mg/kg)、土壤有机碳(soc)(g/kg)、砂粒含量(%)、粉粒含量(%)和粘土含量(%)。
2.5.地上生物量碳
在缺乏区域异速生长模型的情况下,根据先前的研究(Mensahetal,2020c),我们采用Chaveetal(2014)开发的多物种异速生长生物量方程来确定样地中所有单株树木的生物量。多物种异速生物量方程的公式为:
2.6.性状相关多样性及群落特征指数
对于每个选定的功能性状(即Hm,SLA,LDMC和SDM),我们计算了样地水平群落加权平均值(CWM);Garnieretal.,2004)。CWM反映了群落间平均性状值和物种优势度的功能变化。CWM计算为群落的平均性状值,并以物种丰度为权重。在我们的数据集中,单个树的高度和Hm之间的关系的Pearsonrsquared很低(R2=0.17),表明这两个高度在很大程度上是解耦的。
为了量化功能多样性(FD),我们在地块水平上计算了单变量和多变量性状功能分散度(FDiss)。FDiss是一种广泛使用的差异度量(Lalibert'eetal,2022),它衡量了群落内物种之间特征值的变化,同时考虑了它们的相对丰度(Lalibert'eetal,2022)。我们使用FDiss是因为它相对于FD的其他度量具有相当大的优势。首先,它对物种丰富度不敏感,不受异常值的强烈影响。其次,与其他FD指标不同,对于只有一个物种的群落,FDiss的值为0(Lalibertetal,2022)。利用FD软件包中的物种功能性状值和plot-species丰度矩阵计算CWM以及单变量--性状FDiss和多变量--性状FDiss(Lalibert'eetal,2022)。功能性状的属性变化见在线图A5。
2.7.系统发育树
通过将这些物种与软件包中实现的大系统发育联系起来,我们构建了所有物种的扎根和时间校准的系统发育。
2.8.系统发育多样性指标
从系统发育树中,我们计算了两种评估树木群落进化史的系统发育多样性指标:平均成对系统发育距离(MPD)和平均最近分类单元距离(MNTD)(CoelhodeSouzaetal,2019;Tuckeretal,2017)。MPD是物种之间以分支长度表示的平均成对系统发育距离,并被计算为群落中所有成对个体组合之间的平均系统发育距离(Webbetal,2002)。MNTD是个体与最近亲缘(非同种)个体之间的平均距离(Webbetal.,2002)。我们使用系统发育树和picante包中的plot-speciesabundance数据来计算这些系统发育指标(Kembeletal,2010)。我们更倾向于使用MPD和MNTD,而不是其他几种系统发育指标,因为本研究所解决问题的性质,也因为它们是互补指标,对不同系统发育深度的过程很敏感(Mazeletal,2016;Tuckeretal,2017)。通过计算MPD和MNTD的标准化效应大小(ses),我们计算了物种丰富度与MPD和MNTD的潜在相关性(Mazeletal,2016):
式中,Xobs为系统发育多样性度量的观测值;X为系统发育多样性指标随机值的平均值;sd为系统发育多样性指标随机值的标准差。
2.9.统计分析
采用R统计软件进行统计分析。为了确定在基于功能性状的生态模型中纳入系统发育信息的重要性,我们使用了线性混合效应模型(lmm)和部分冗余分析(pRDA)。具体来说,我们拟合了三个lmm,其中地块大小(plotsize)作为随机因素,AGC作为响应变量,其中我们分别测试了(i)PD指标(sesMPD和sesMNTD)的影响;(ii)功能性状(FDiss和CWM)的生物多样性属性效应;(iii)PD、FDiss和CWM的综合效应。根据Nakagawa和Schielzeth(2013)计算了边际R平方和条件R平方。通过检查(i)模型残差的正态性,(ii)预测变量之间的相关性(图A6)和(iii)通过方差膨胀因子进行多重共线性,对lmm进行了补充分析。在pRDA中,我们保持效应区大小不变,并确定了AGC的变化有多少可以由(i)PD,(ii)FDiss和CWM以及(iii)PD,FDiss和CWM解释。进行排列试验以检验pRDA的整体意义。最后,采用方差分区分析方法,根据PD和功能性状属性对唯一方差和共有方差进行划分。lmm使用lme4包进行(Batesetal,2015),而pRDA、排列测试和方差划分分析使用vegan包进行(Oksanenetal,2022)。
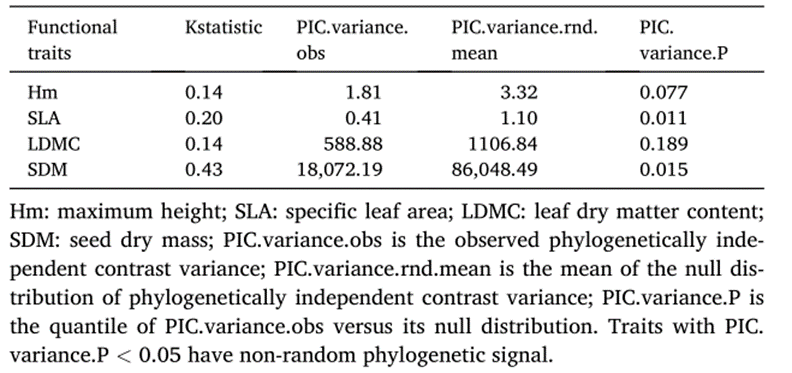
为了了解PD与FDiss和CWM的亲缘关系沿系统发育信号梯度的轨迹,我们首先使用Blomberg的K统计量(Blombergetal.,2003)测试了每个功能性状的系统发育信号。接下来,我们分析了PD指标(sesMPD和sesMNTD)对各功能性状的FDiss和CWM的双变量影响。这些双变量效应是从16个独立的LMM(以图大小作为随机因子)的结果中提取的,测试PD指标对四个功能性状中每一个的FDiss和CWM的个体影响(见表A1-A2)。为了更好地描述pd--性状关系的趋势,我们通过拟合lmm,重新模拟了sesMPD和sesMNTD对每个功能性状的FDiss和CWM的个体影响,同时允许截距和斜率随地块大小而变化。然后,使用merTools包中的REsim函数(Jaredetal,2023)基于lmm结果的后验分布来模拟这些效应。最后,我们根据sesMPD和sesMNTD对每个性状的FDiss和CWM的标准化效应的大小,绘制了bloombergK统计量作为系统发育信号的梯度。
为了分析系统发育和功能性状属性之间的相互关联路径是否以及如何联系AGC和环境变量,我们基于图1b中的假设和概念模型拟合了一个分段结构方程模型(pSEM),并通过FDiss和CWM的变化检验PD对AGC的影响。我们通过使用最重要的AGC环境预测因子来简化pSEM框架,这些预测因子是通过交叉验证优化确定的,该优化是使用gbm包的增强回归树(BRT)分析进行的(Greenwelletal,2022)。对于FDiss,我们使用了前面定义的多性状指数,而对于CWM,我们使用了CWMHm,它被认为是AGC的重要预测因子。基于方向分离试验(d-separationtest)(Shipley,2013)进行验证性路径分析,判断模型是否需要引入缺失路径。使用Fisher的C统计量和相关的p值来评估pSEM的拟合性(Shipley,2013)。使用分段式esem软件包安装pSEM(Lefcheck,2016)。
3.结果
3.1系统发育多样性对性状模型的影响
根据部分冗余分析(保留样块大小异质性不变的影响)和排列检验,基于功能性状的模型具有统计学意义(p=0.001),解释了11.01%的AGC方差,而基于系统发育多样性(sesMPD和sesMNTD)的模型(也具有全球显著性,p=0.023)仅能解释了1.95%的AGC变异(表1)。这表明PD对AGC变异的解释低于功能性状。在解释AGC变异方面,功能性状相对于PD更为重要。
表1
部分冗余分析(pRDA)和排列测试的结果,
以及测试性状和系统发育多样性对AGC影响的线性混合效应模型的结果
当我们将功能性状属性和系统发育多样性结合在同一模型中,只有CWMHm()和sesMPD(p=0.023)对AGC有显著影响(表1)。尽管叶片干物质含量功能分散(FDissLDMC)、种子干质量功能分散(FDissSDM)和多性状功能分散(FDiss)等单个性状指标与AGC有显著相关性(图A6)。在综合多重模型中,它们对AGC的影响不如CWMHm和sesMPD重要(表1)。将系统发育信息纳入基于性状的模型后,解释方差增加到12.3%(即15%)。线性混合效应模型也观察到了类似的趋势(表1)。这些发现表明,在理解PD、FDiss和CWM对AGC的重要性方面,综合模型比单个模型更具优势。
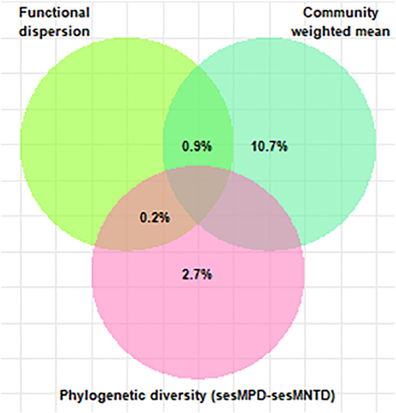
方差划分分析(图3)表明,性状的CWM和系统发育多样性(sesMPD和sesMNTD)分别解释了10.7%和2.7%的AGC的独特差异,而AGC中只有0.2%的差异可以同时解释系统发育多样性和功能单性状多样性。
图3
韦恩图说明了通过单性状FDis(功能分散)、群落加权平均值(CWM)和系统发育多样性解释的地上碳的共享和独特方差
3.2.沿系统发育信号梯度的系统发育-性状关系
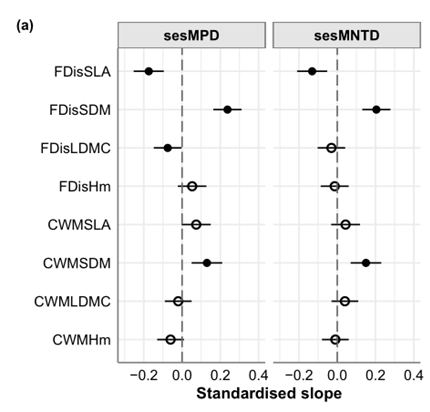
在性状的系统发育信号检测中,我们发现SDM和SLA表现出显著的信号(SDM:Blomberg’sK=0.43,p=0.015;SLA:Blomberg’sK=0.20,p=0.011),表明这些功能性状比Hm和LDMC更具有系统发育保守性(表2)。系统发育多样性与单个性状功能指标之间的双变量关系的标准化斜率对于系统发育保守性状(即SLA和SDM)中通常是显著的,而Hm和LDMC则不显著(图4a;表a1a2)。此外,FDis对这些斜率的影响大于CWM。与CWM不同的是,FDis的斜率随系统发育信号的增加而增加,这表明CWM对系统发育保守性的响应不如FDis(图4b)。
表2
本研究中使用的功能性状和测试其系统发育保守性的Blomberg’sK统计量
图4
(a)
系统发育多样性与单一性状功能指数之间的关系
图4(b)
系统发育多样性-性状关系的斜率大小沿系统发育信号梯度的轨迹
3.3.性状及其与AGC和环境的系统发育关系
基于BRT分析(图A8),我们将气候湿度指数和土壤全氮作为AGC最重要的环境预测因子,与系统发育多样性指标(sesMPD和sesMNTD)和多性状功能分散(FDiss)一起纳入pSEM中。pSEM与数据拟合良好(p=0.419;图5),并表明sesMPD(β=0.24)和CWMHm(β=0.31)的增加与AGC储量的增加相关(图5;表A3)。FDis与AGC无显著相关,但与sesMPD呈正相关(β=0.23;p=0.016),表明增加sesMPD可能会增加互补效应的机会。CWMHm在系统发育多样性指标上没有表现出显著的变化路径,同时受土壤肥力和气候湿度指数的共同影响。总体而言,pSEM结果表明,sesMPD和CWMHm在预测AGC与土壤肥力和气候湿度指数之间的关系方面优于FDis。
图5
分段SEM总结,评估系统发育多样性和功能性状属性之间的相互关联路径,及其与环境变量(土壤和气候)和地上碳的联系
箭头是假设的因果路径。路径旁边的值是标准化路径系数,表A3中给出了其重要性的完整细节。R2m:边缘R平方;R2c:条件R平方;黑色素线表示重要路径,灰色和虚线表示不显着影响。
4.讨论
4.1.系统发育多样性对解释AGC变异很重要,即使在考虑了功能性状之后也是如此
4.2.系统发育-性状关系对FDis性状保守性比CWM更敏感
4.3.环境因子通过系统发育多样性和功能性状间接影响AGC
总而言之,本研究为我们理解热带半干旱区树木系统发育和性状的影响及其与环境和生态系统功能(如地上树木碳储量)的关系具有重要意义。这表明,系统发育多样性在解释地上部木材碳多样方面很重要,但不是功能性状。我们的模型解释了较低的AGC方差,这表明其他预测因子,如林分结构变量,可能是影响半干旱植物群落树木生物量碳的额外决定因素。在我们的研究中,性状与系统发育之间的相互关系比群落加权平均值更具有响应性,并且性状保守性对功能多样性的影响更大。最后,气候湿度和土壤氮等环境因子通过对系统发育多样性和功能性状的影响间接影响地上碳。本研究结果反映了(i)环境过滤导致的性状趋同(如优势树种对弱势树种的竞争性排斥),以及(ii)在干旱程度较低的环境中近亲物种的进化过程得以维持。这些过程强调了系统发育多样性和群落性状在调节森林和热带稀树草原地上碳储量中的作用。
SylvanusMensaha,b,*,Kangb́eniDimobec,FlorentNoul`ekound,FonsvanderPlase,ThomasSeiferta,Phylogeneticdiversityandcommunitywide-traitmeansofferdi(2024)167905
免责声明:本文章如果文章侵权,请联系我们处理,本站仅提供信息存储空间服务如因作品内容、版权和其他问题请于本站联系